Биомеханика – это фундаментальный процесс, который включает в себя создание силы мышечными волокнами. Исследования этих принципов механических свойств применительно к биологическим системам невероятно интересны. Представленный обзор публикаций демонстрирует недавние исследования, в которых используется оборудование Aurora Scientific для достижения успехов в области биомеханики.
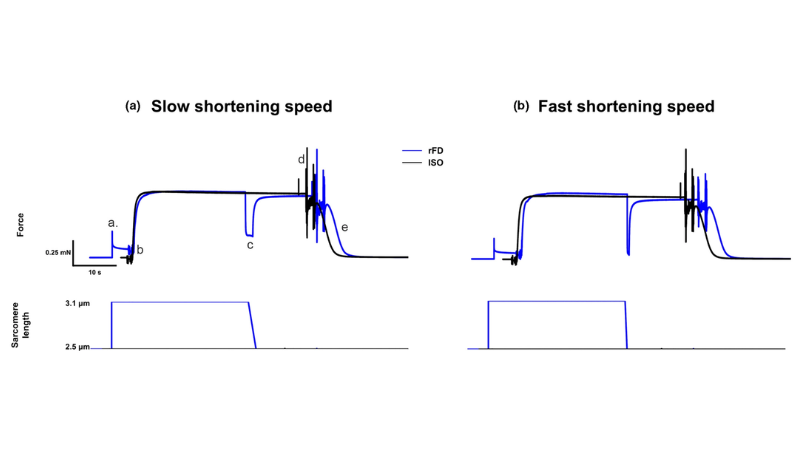
На изображении (©Mashouri et al. (2021)) показаны графики силы для условий медленного и быстрого активного сокращения в зависимости от подавления остаточной силы.
Влияние скорости сокращения на соотношение жесткости и силы при восстановлении изометрической силы предполагает механизмы снижения остаточной силы
В статье Jeong & Nishikawa (2023) исследуется феномен подавления остаточной силы, который относится к уменьшению стационарной изометрической силы после активного сокращения и восстановления силы. В исследовании изучается соотношение мышечной жесткости (km) и изометрической силы (FISO) в период восстановления изометрической силы после активного сокращения на разных скоростях. Предыдущие исследования измеряли это соотношение в один момент времени, в то время как данное исследование направлено на измерение его в разное время после начала восстановления изометрической силы. Авторы предположили, что отношение km к FISO может дать представление о механизмах, лежащих в основе подавления остаточной силы.
В дизайне исследования мышцы развивают силу изометрически с последующим сокращением с разной скоростью и различной продолжительностью восстановления изометрической силы. Авторы использовали двухрежимный рычаг 300B для проведения этих измерений на камбаловидных мышцах мыши. Таким образом, исследование показало, что на соотношение жесткости и силы во время восстановления изометрической силы влияет скорость активного укорочения. Хотя слабосвязанные поперечные мостики могут способствовать увеличению жесткости, следует также учитывать и другие механизмы, такие как восстановление искажений актина или участие титина в качестве настраиваемого эластичного элемента. Эти результаты способствуют лучшему пониманию механизмов, лежащих в основе подавления остаточной силы.
Увеличение силы после растяжения волокон диафрагмы и миофибрилл сопровождается увеличением неоднородности длины саркомера и чувствительности к Са2+
В статье Contini et al. (2022) обсуждают феномен увеличения остаточной силы (RFE) в скелетных мышцах, который относится к увеличению и стабилизации силы, когда мышечное волокно растягивается во время активации. В исследовании подчеркивается, что RFE не наблюдается в сердечных мышцах. Присутствие RFE в скелетных мышцах объясняется сочетанием статического напряжения и неравномерностью длины саркомеров во время активации и растяжения. Статическое напряжение связано с увеличением жесткости белка титина. В статье также отмечается отсутствие исследований по изучению RFE в мышце диафрагмы, которая обладает уникальными характеристиками, обусловленными ее ролью дыхательной мышцы.
В ходе исследования были проведены эксперименты с использованием интактных живых волокон и изолированных миофибрилл мышей для оценки наличия и механизма RFE в диафрагме. Авторы подготовили образцы мышц, используя аппарат для малых интактных мышц 801C Aurora Scientific, высокоскоростной контроллер длины 322C и датчик силы 405A. Они стимулировали мышечные волокна с помощью электрического симулятора 701C Aurora Scientific и анализировали полученные данные с помощью системы сбора и анализа данных о мышцах в реальном времени 600A. Авторы наблюдали RFE в диафрагме, что сопровождалось увеличением неравномерности длины саркомеров и чувствительностью миофибриллярного кальция к кальцию. Затем они смогли понять, как неоднородность длины саркомеров и повышенная жесткость тайтина в миофибриллах могут привести к RFE, построив модель, которая смогла воспроизвести экспериментальные наблюдения.
Модификация подавления остаточной силы в отдельных мышечных волокнах после тренировок крыс на подъем и спуск
В статье Mashouri et al. (2021) ученые исследует подавление остаточной силы (rFD), уменьшение силы после сокращения мышц. В исследовании изучается возможность модификации RFD на уровне отдельных мышечных волокон и ее связь с типом волокон и тренировкой. Предыдущие исследования показывают, что на RFD может влиять объем работы, выполняемой во время укорочения. Тип волокна оказал некоторое влияние на RFD, но результаты неубедительны. Тренировочные исследования не обнаружили последовательных изменений RFD на уровне всей мышцы. Целью исследования является определить, можно ли модифицировать RFD путем тренировки крыс на подъем и спуск.
Авторы взяли крыс и подвергли их бегу вверх или вниз в течение 4 недель, после чего исследовали их мышцы-разгибатели пальцев (EDL) и камбаловидную мышцу (SOL). Используя датчик силы 403A Aurora Scientific и высокоскоростной контроллер длины 322C, длину саркомеров отдельных волокон устанавливали с помощью высокоскоростной видеосистемы 901D для определения длины саркомера. Результаты показывают, что RFD присутствовал в обеих мышцах, без каких-либо изменений после тренировки. Мышца EDL демонстрировала большее подавление RFD и жесткости по сравнению с мышцей SOL. Эти данные свидетельствуют о том, что RFD является внутренним свойством отдельных мышечных волокон, которое не поддается изменению тренировками.

















